Els bacteris són un conjunt d'éssers vius procariotes unicel·lulars que només són visibles al microscopi i que existeixen a la Terra des de fa més de 3 mil milions d'anys. Amb el transcurs del temps, aquests éssers han anat evolucionant fins arribar a un punt en què la seua presència és sinònim de l'aparició de diverses malalties en els organismes als que infecten, guanyant-se la qualificació d'agents patògens. Entre els organismes que es veuen afectats ens trobem nosaltres, els éssers humans. No obstant això, la nostra intel·ligència i curiositat ens ha permès idear -més aviat descobrir- determinades substàncies per lluitar contra aquestes cèl·lules, els antibiòtics. Tot i l'eficàcia que posseeixen actualment, cada vegada hi ha més antibiòtics que deixen de ser útils per a combatre els bacteris, ja que han aparegut uns peculiars bacteris que són capaços de sobreviure a ells, els anomenats superbacteris. En aquest treball es tractarà d'explicar el terme de superbacteri, junt amb les diferències—genètiques— que les separen dels bacteris corrents i la manera que tenen de transferir-les tant a altres bacteris com a la seua descendència. A més a més, es tractarà la relació històrica que hi ha hagut entre els superbacteris i els antibiòtics, explicant així l'efecte de l'ús irresponsable d'aquests últims sobre els primers. Així mateix, s'exposaran les conseqüències que pot comportar l'augment de la població d'aquests bacteris; així com els mètodes per prevenir i combatre el seu creixement, esmentant també moviments o mètodes actuals que tenen per objectiu eliminar aquests microorganismes.
1. Superbacteris
1.1. Definició i origen del mot
El terme "Superbacteris" (1) va sorgir, segons les fonts de LexisNexis, a la premsa habitual dels anys 70, i el feia servir per referir-se a bacteris eliminadors de contaminants.
Actualment, el malnom "superbacteri" es refereix a aquells microorganismes dotats de múltiples gens que proporcionen alts nivells de resistència als antibiòtics específicament recomanats per al seu tractament, implicant una major morbiditat i mortalitat. L’adquisició de resistència antibiòtica pot produir-se a partir de l’aparició de mutacions genètiques que modifiquen els gens i, per tant, la informació necessària per a la síntesi de polipèptids, donant lloc a molècules que poden tenir un efecte inhibidor en les substàncies (2) que, en un principi, indueixen a la lisi cel·lular. L'origen d'aquests canvis genòmics(1, 3, 4) -i per tant de la resistència antibiòtica- es deu a l'ús excessiu dels mateixos antibiòtics; acció que sotmet a les poblacions bacterianes a una gran pressió selectiva on, segons el mecanisme de selecció natural, les que sofreixen mutacions que porten al desenvolupament de caràcters més avantatjosos— és a dir, les que desenvolupen resistència cap a l’antibiòtic— seran les que sobrevivisquen, deixant de veure’s afectades pel fàrmac.
Aquesta situació, ja prou perillosa per si mateixa, es va veure agreujada per la inesperada identificació de la resistència antibiòtica genèticament transferible a Japó, a mitjans de la dècada de 1950, ja que, a part de la selecció natural, també s'havia de tenir en compte que els gens que confereixen resistència a antibiòtics es podien transmetre horitzontalment— transmissió del DNA dels plasmidis, o fragments de DNA, a altra cèl·lula—entre tota una població de bacteris patògens, facilitant així la seva dispersió (4).
El descobriment dels superbacteris es remunta a mitjans del segle XX, poc després de l'aparició de la penicil·lina. La penicil·lina és un antibiòtic, descobert per Alexander Fleming (Figura 1) al 1928, que va revolucionar la indústria farmacèutica per la seva capacitat de provocar la lisi de bacteris causants de diverses malalties. L’any 1940, diversos anys abans de la introducció de la penicil·lina com a agent terapèutic, es van identificats bacteris amb un enzim anomenat penicil·linasa, amb la capacitat d’hidrolitzar i inactivar la penicil·lina, encara que en la comunitat científica no se li va atorgar la importància requerida en eixe moment. No obstant això, després d'haver fet servir l’antibiòtic àmplia i excessivament, l'aparició de soques amb penicil·linases capaces d’inactivar-lo va arribar a ser bastant freqüent, havent-se de dur a terme estudis sintètics en resposta per tal de modificar químicament la penicil·lina i prevenir la seva escissió per l’acció d'aquests enzims. Aquests estudis van desembocar en la invenció de la meticil·lina al 1959, un antibiòtic capaç de combatre contra les soques resistents a la penicil·lina. Posteriorment, però, va sorgir la MRSA (methicillin-resistant S. aureus), resistent a aquest antibiòtic (3).
Figura 1.- Alexander Fleming al seu laboratori.
Existeixen bastants casos similars al de la penicil·lina, com el de l'estreptomicina, introduïda al 1944 com a tractament per a la tuberculosi; va provocar la proliferació de soques mutants de Mycobacterium tuberculosis resistents a l'antibiòtic (4). De fet, aquest bacteri, juntament amb molts dels patògens bacterians associats amb les epidèmies, ha evolucionat fins a donar lloc a formes multiresistents a diversos antibiòtics.
Un d’aquests és l’anomenat S. aureus (patogen Gram-positiu), del qual després d’adquirir resistència a la meticil·lina van sorgir variants multiresistents del mateix bacteri (un total de 6 espècies, totes amb diferents combinacions de gens de resistència). Actualment, és la infecció nosocomial principal; fa relativament pocs anys, el bacteri s’ha propagat fora de l’hospital i ara és considerat un dels principals patògens adquirits per la població no hospitalitzada (Community-associated MRSA infections, CA-MRSA), amb una virulència i una transmissió augmentada i amb nous grups de gens de patogenicitat (5), com el gen que codifica per a la toxina citolítica leucocidina de Panton-Valentine. També és interesant l’adquisició del gen vanA, codificat en un transposó derivat del genoma de Enterococcus spp., el qual segurament va adquirir per transferència horitzontal i que confereix resistència a la vancomicina, antibiòtic utilitzat per tractar les infeccions de MRSA.
Altres casos de patògens nosocomials són Pseudomonas aeruginosa, de gran preocupació per als pacients amb fibrosi quística, ja que el seu desenvolupament de resistència està associat amb el tractament antibiòtic prolongat de pacients amb aquesta malaltia. És un superbacteri molt persistent i pot evadir les defenses immunitàries humanes (2). Altres, com Acinetobacter baumannii, un patogen Gram-negatiu més recent, posseeix un conjunt de gens de resistència i determinants de patogenicitat que resulten en l’augment de taxes de la mortalitat i la morbiditat.
Realment, no està del tot clar què fou abans, la resistència o l'antibiòtic; ja que, per exemple, la penicil·linasa es va descobrir abans de l'ús de la penicil·lina com a tractament terapèutic (6). No obstant això, una cosa està clara, i és que l'excessiu ús d'antibiòtics és el causant de la ràpida proliferació de les soques resistents.
1.2. Motius i mecanismes d’adquisició de resistència
Fins fa relativament poc, contraure una xicoteta infecció podria causar la mort a qualsevol persona jove i sana. Sotmetre’s a una operació quirúrgica, anar al dentista, fer-se una ferida... resultava tota una loteria que moltes vegades acabava en tragèdia. Fins que això canvià amb l’arribada dels antibiòtics. La qualitat de vida de la població i, en conseqüència, l’esperança de vida mitjana d’aquesta augmentà en pocs anys.
No obstant això, els antibiòtics han perdut efectivitat progressivament i ja ho va dir Alexander Fleming, descobridor dels efectes antibiòtics de la penicil·lina, durant el discurs que oferí el 1945 en guanyar el premi Nobel de Fisiologia i Medicina: “ Arribarà el dia en què la penicil·lina podrà ser comprada en qualsevol de les botigues. Llavors existirà el perill de que un home ignorant puga fàcilment prendre una dosi insuficient i, en exposar els bacteris a quantitats no letals del fàrmac, els faça així resistents." Set dècades després, el presagi es compleix.
1.2.1. Motius de resistència
¿De quina manera s’ha arribat a parlar de “superbacteris” com a una amenaça per a la salut mundial? Cal tindre en compte dos factors per respondre a aquesta pregunta: el social i el mèdic.
Segons s’ha valorat, un 50% dels antibiòtics emprats en els Estats Units són inapropiats, perquè han estat receptats per a infeccions virals, en dosis inadequades o en un lapse de temps massa inapropiat. D’aquesta manera, nous antibiòtics eren receptats, arribant així a un punt en què el nombre de nous antibiòtics receptats era massa gran, comparat amb el nombre de nous antibiòtics desenvolupats.
Una de les fonts més importants de resistència en bacteris, és l’ús excessiu d’antibiòtics en el ramat de granges i corrals industrials per millorar el seu creixement. Més tard, aquests bacteris resistents passen als humans que ingereixen la carn del ramat; procés conegut com a zoonosi.
Altra font mèdica-social de resistència a antibiòtics són els propis hospitals, on la propagació dels bacteris és molt senzilla. Processos com els trasplantaments, immunodepressions o l’ús d’implants (afavoreixen el creixement de biofilms) són destacables a l’hora d’establir els orígens actuals dels superbacteris.
El fet d’adquirir resistència a antibiòtics resulta, en tota comunitat bacteriana, una pressió de selecció molt gran, perquè sols eixe bacteri resistent, o eixos pocs, podran transmetre a la seua descendència, o a altres bacteris mitjançant transferència horitzontal, els gens de resistència. La plasticitat genètica present en bacteris, a causa de la gran capacitat de transmetre fragments de DNA i el poder d’assimilar-los i fer-los propis, és un altre factor a tindre en compte pel que fa als motius de resistència.
És a dir, el mal ús dels antibiòtics, l’excés d’aquests i la irresponsabilitat de tota la societat, han provocat que els bacteris, éssers molt temuts, hagen “pujat de nivell”, fins arribar a tal punt que les armes definitives contra ells no siguen eficients.
Una vegada vistes les causes per les quals s’adquireix resistència, ¿com fan els bacteris per resistir contra productes, a priori, molt nocius per a ells?
Els mecanismes de resistència es poden agrupar en quatre grans ordres, segons la seua manera general d’operar:
a) Limitant l’accés dels antibiòtics.
b) Inactivant l’antibiòtic mitjançant enzims.
c) Modificació o protecció de l’objectiu de l’antibiòtic.
d) Regulació dels gens de resistència.
Cal remarcar que, tot i que fem una classificació dels mecanismes per separat, els Superbacteris poden combinar distints efectes per augmentar el seu escut contra els antibiòtics. Per exemple, els MDR (multidrug resistant) tenen diversos mecanismes contra una gamma d’antibiòtics considerable.
a) Limitant l’accés dels antibiòtics. Una manera d’impedir que els antibiòtics entren en contacte amb el bacteri és presentant una membrana externa (a la plasmàtica) que dificulte l’arribada de les substàncies nocives a la membrana plasmàtica. Per exemple, els β-lactams, com la penicil·lina, han de travessar la membrana externa dels gram-negatius per arribar al seu lloc d’unió en la membrana plasmàtica. Òbviament, existeixen a la membrana externa porines perquè els nutrients del medi puguen difondre i arribar als bacteris. Tanmateix, l’expressió d’unes proteïnes o altres pot ser regulada de manera que la permeabilitat de la membrana pot augmentar o disminuir. Així, en cas d’antibiòtic en el medi, un bacteri augmentarà l’expressió de porines que formen barrilets β més xicotets, per on els antibiòtics no puguen passar. O, per altra banda, secretarà proteïnes que formen porus més grans per on antibiòtics voluminosos difonguen, i així sensibilitzar-se més a aquests compostos. En Pseudomonas aeruginosa hi ha més de 70 gens, dels 5500 presents al seu genoma, que codifiquen per a proteïnes de porus.
Altra forma efectiva d’impedir l’accés de l’antibiòtic al citoplasma és amb una bomba activa. És a dir, proteïnes de membrana que bomben (traslladen amb un cost energètic) substàncies a un lloc concret de la cèl·lula -o fora d’aquesta- en contra de gradient, poden ser usades pels bacteris per evitar una alta concentració d’antibiòtic dins d’ells, assolint així l’estat resistent. Normalment, aquestes bombes tenen un paper corrent en les cèl·lules com és mantenir l’homeòstasi de l’individu expulsant metabòlits secundaris o altres substàncies de rebuig. Així doncs, els bacteris poden tenir moltes bombes al llarg de la seua membrana que siguen específiques per a certes substàncies o que abasten un rang ampli d’aquestes. D’aquesta forma poden arribar a ser resistents a molts antibiòtics. Aquest és un mecanisme de resistència, recentment descobert, que està resultant problemàtic tant per a metges com per a investigadors.
b) Inactivació enzimàtica de l’antibiòtic. Les β-lactamases són enzims que hidrolitzen l’anell dels antibiòtics anomenats β-lactams, deixant-los inactius. El mecanisme d’hidròlisi és mostra a la Figura 2.
Figura 2.- La serina de les B-lactamases que contenen aquest aminoàcid, forma un enllaç covalent amb l'anell dels B-lactams.
Aquest enllaç permet que les molècules d'aigua l'ataquen, obrint l'anell i inactivant l'antibiòtic (Wilson, BA et al. 2007).
Aquests enzims hidrolítics es poden inactivar mitjançant altres substàncies que impedeixen la unió de la serina del lloc d'unió a l’anell. El problema és que per a cada inhibidor de β-lactamases que es troba, sorgeix una nova β-lactamasa a la que eixos inhibidors no afecten. Aquest fet es deu a què aquestes hidrolases són molt específiques i coevolucionen ràpid amb els seus inhibidors. A més, tots els inhibidors de β-lactamases que s’han sintetitzat o descobert, sols inhibeixen un tipus d’aquestes, les associades a serina, ja que n’hi ha d’altres amb un cofactor metàl·lic que no es poden desactivar artificialment.
Un altre mecanisme característic dels bacteris resistents, és l’oxidació de les tetraciclines, una mena d’antibiòtics. L’enzim que oxida i inactiva aquests antibiòtics, necessita O₂ i NADPH per dur a terme la reacció, i està codificat pel gen tetX.
c) Modificació o protecció de l’objectiu de l’antibiòtic. Dins d’aquest tipus de mecanismes podem trobar dos subtipus.
El primer d’ells és l’acumulació de mutacions per la forta pressió de selecció que produeixen els antibiòtics; sobre tot, els bacteriostàtics que no maten els bacteris físicament. Un exemple clar d’aquest mecanisme es troba en el residu 2058 de la subunitat 23S del rRNA dels bacteris (una adenina). Aquest residu d’adenina està relacionat amb els ponts d’hidrogen que estableixen certs antibiòtics amb aquesta subunitat de RNA ribosòmic, impedint així que esdevinga el procés de traducció. Quan aquesta adenina muta a guanina, eixos ponts d’ hidrogen es perden, i llavors apareixen els fenòmens de resistència.
El segon mecanisme és la modificació química de les dianes dels antibiòtics, per impedir la unió amb aquests, però de manera que la funció inicial de la diana no es vega afectada. Per exemple, la vancomicina inhibeix l’entrecreuament dels peptidoglicans unint-se als seus residus finals D-Ala-D-Ala com es veu a la Figura 3.
Figura 3.- Mecanisme d'acció de la vancomicina en bacteris sense resistència. (Wilson, BA et al. 2007)
En principi, un canvi en la seqüència D-Ala-D-Ala fou difícil d’aconseguir, però la vancomicina s’emprà molt àmpliament durant un període de temps suficient per tal que els bacteris trobaren la forma de resistir: canviant la seqüència a D-Ala-D-lactat. Per a açò, són necessaris tres enzims que catalitzen diverses reaccions: la fermentació de piruvat a lactat, la síntesi de la seqüència D-Ala-D-lactat i la hidròlisi de la seqüència que se sintetitza per via normal, respectivament. De manera que, la vancomicina no pot unir-se encara que la capacitat d’entrecreuar els peptidoglicans segueix present (Figura 4).
Figura 4.- Mecanisme bacterià per resistir a la vancomicina. (Wilson, BA et al. 2007).
d) Regulació dels gens de resistència. Els gens que confereixen resistència a antibiòtics per part dels bacteris no són gens que produeixen el seu producte gènic constantment; és a dir, no són constitutius, perquè sols s’han d’expressar en presència d’antibiòtics. Llavors, és lògic pensar que la seua expressió haurà d’estar regulada com la resta de gens regulats.
Els processos normals de regulació gènica estan presents en aquests mecanismes: la repressió, l’activació i el control de la traducció són iguals ací que amb la resta de gens.
Figura 5.- Transferència d'un transposó (Wilson, BA, 2007).
No totes aquestes estratègies desenvolupades pels bacteris de resistència als antibiòtics són exitoses en produir-se un sol canvi en el genoma. Els al·lels dels gens que confereixen resistència apareixen per mutació, però no sempre amb la primera mutació apareix el mecanisme de resistència. És un fet recurrent que per obtenir resistència no siga necessària una sola mutació, sinó una acumulació de moltes d’elles, fent així més lent i més complicat el procés. Per tant, el cost de les poblacions bacterianes per trobar una resistència eficaç contra algun antibiòtic és ben alt i no lliure de risc.
Una manera, molt més ràpida d’adquirir resistència a antibiòtics, és obtenint els gens que aporten aquesta resistència mitjançant transferència horitzontal de gens (THG). S’ha comprovat que aquest fenomen és molt freqüent entre bacteris en la natura. I, filant més prim, es poden transferir plasmidis amb més d’un gen de resistència; és a dir, es pot conferir resistència a més d’un antibiòtic amb un sol procés de transferència horitzontal.
Dels tres processos possibles de transferència horitzontal, el més reiteratiu és la conjugació, perquè no té restriccions com les altres dues (la transducció, per exemple, necessita que el bacteri tinga un receptor concret per al bacteriòfag).
No obstant això, no tots els gens de resistència a antibiòtics se situen en plasmidis, ni tots els plasmidis són capaços de transferir-se. Els plasmidis que tenen més d’un gen de resistència, poden haver obtingut aquests mitjançant transposons de conjugació. Aquests funcionen com a transposons normals, però amb la capacitat afegida de poder transferir-se del genoma d’una cèl·lula al d’altra cèl·lula, com es veu a la Figura 5.
D’aquesta forma, el transposó provinent d’altre bacteri pot transferir-se, més tard, a un plasmidi amb altre gen de resistència, assolint així la característica de bacteri multiresistent.
Per tant, el fet d’existir-hi bacteris resistents a antibiòtics, no és preocupant tan sols pel fet que siguen molt difícils d’eliminar, sinó també pel fet que la resistència es pot transmetre fàcilment, produint milers de bacteris resistents.
1.3. Prevenció contra la infecció i l’emergència de noves resistències antibiòtiques
1.3.1. La higiene
A mitjans del segle XIX, la mort d’una dona durant o poc després del part a causa de l’anomenada febre puerperal (7) era una situació natural. Aquesta malaltia no es considerava més que una conseqüència nefasta i moltes vegades inevitable del fenomen de donar a llum. No va ser fins els febrer de 1846 quan la resignació sobre aquest assumpte vegué un aguait d’esperança.
Figura 6.- Retrat d'Ignác Fülöp Semmelweis (chronotopeblog.com)
En aquests temps, el jove Ignác Fülöp Semmelweis (Figura 6) es trobava treballant com a ajudant en la secció d’obstetrícia de l’Hospital General de Viena. Semmelweis començà a investigar la possible causa d’aquest fet i, sospitant que la malaltia es podria transmetre pel contacte de les mans dels metges a les pacients, fixà aquest cartell a l’hospital:
“A partir de hui, tot metge o estudiant que vaja des de la sala d’autòpsies cap a la de parts, està obligat a rentar-se acuradament les mans en una palangana amb aigua daurada disposada en la porta d’entrada.” A finals d’any, la mortalitat en la secció de Semmelweis baixà a un 12%. Un mes més tard, en ampliar la norma d’higiene per a totes les situacions d’exploracions a pacients; la xifra baixà a 0,23%.
Aquesta xicoteta història reflexa a la perfecció la importància de la higiene. Hui en dia sembla impensable que es torne a donar una situació semblant a l’anterior, no obstant, existeixen moltes altres situacions en què es pot estar exposat a infeccions sense ser-ne conscients. El primer pas és la precaució per evitar possibles infeccions bacterianes i, més concretament, aquelles potencialment perilloses causades pels superbacteris.
Una bona higiene de mans (8), com descriu la història anterior, pot ser clau per evitar malalties infeccioses. És per això que l’OMS (Organització Mundial de la Salud) i l’ Aliança Mundial per a la Seguretat del Pacient han redactat guies d’aplicació global per a la higiene de mans (9), en la qual es detallen els productes més adequats per a la neteja de mans i ungles, com s’han d’usar, el moment en què és indicat aplicar-los, etc. (Mireu en la Figura 7 un exemple de com podem tindre les mans, alguns de nosaltres, encara que no ho semble).
Figura 7.- Fotografia guanyadora del primer premi de la 13a edició del seu certament Fotociencia. Empremta d'una xiqueta de 6 anys després de posar la seua mà sobre una placa de Petri.
Es poden destacar l’aplicació de productes de neteja específics com preparats de base alcohòlica que inactiven els microorganismes o detenen la seva proliferació temporàniament, i rentar-se sempre les mans amb sabó, ja duguen productes antisèptics o no.
D’altra banda, és també molt aconsellable:
- mantenir una bona cura odontològica per previndre les càries, l’halitosi, la gingivitis o la periodontitis (10).
- mantenir relacions sexuals de manera segura per evitar la contracció de malalties de transmissió sexual bacterianes com la clamídia (Chlamydia trachomatis), la gonorrea (N. gonorrhoeae) o la sífilis (Treponema pallidum).
- tossir i esternudar sempre que siga possible en mocadors de paper i després llançar-los.
- tindre especial cura de les ferides obertes.
- mantenir els animals de companyia nets, desparasitats i vacunats.
- rentar i conservar els aliments en condicions adequades (11).
En casos en els que no existeix un tractament efectiu per a una infecció, la vacunació podria ser l’única forma de protegir els pacients de malalties greus.
Les vacunes són un preparat d’antígens inactivats (innocus). El mecanisme d’actuació es basa en la memòria que presenta el sistema immunològic en els vertebrats (Figura 8). Quan el preparat d’antígens és inoculat dins un organisme, el sistema immunològic d’aquest reconeix l’agent extern i desencadena tota una mena de reaccions per eliminar-lo alhora que es formen cèl·lules memòria de tots els tipus de limfòcits (T i B) específics del mateix antigen. Les cèl·lules memòria permeten que la reacció immunològica siga més ràpida en un segon contacte inclòs varis anys després del primer.
Figura 8.- Resposta immunològica primària i secundària (epideiologiamolecular.com)
Malauradament, hui en dia encara no existeixen vacunes efectives contra patògens resistents a múltiples drogues, ni tan sols per a les tres amenaces que els Centres per al Control i la Prevenció de Malalties dels Estats Units consideren “urgents”: Clostridium difficile, bacteris de la família Enterobacteriaceae resistents als carbapenemes, i N. gonorrhoeae resistent a les sulfamides, la penicil·lina, les tetracicilines, les quinolones, la ceftriaxona, etc.
La vacunació s’ha d’entendre no sols com un mètode efectiu per a evitar el progrés de les malalties, sinó com una altra forma d’evitar el sorgiment de noves formes de patògens resistents. L’aplicació correcta dels calendaris de vacunació a nivell mundial disminuiria de forma significativa la necessitat de consumir medicaments, incloent-hi els antibiòtics.
1.3.3. Conscienciació i educació
Cada 18 de novembre se celebra el Dia Europeu per a l’Ús Prudent dels Antibiòtics; una iniciativa europea de salut pública anual. El seu objectiu és sensibilitzar sobre l’amenaça que la resistència als antibiòtics suposa per a la salut pública (14).
Durant aquest dia, en diversos països de la Unió Europea entre els que s’inclou Espanya, es porten a terme diferents campanyes de conscienciació que ofereixen tota la informació necessària per tal que la correcta actuació dels ciutadans frene poc a poc el desenvolupament de bacteris resistents i aconseguir que els antibiòtics mantinguen la seva eficàcia.
1.3.4. El paper de l’OMS
En maig del 2015 s’aprovà un pla d’acció mundial. La seva finalitat és assegurar que es puga continuar prevenint i tractant les malalties infecciones mitjançant fàrmacs segurs i eficaços. El pla consta de cinc objectius primordials:
1. Millorar la sensibilització i els coneixements en matèria de resistència als antibiòtics.
2. Reforçar la investigació en matèries relacionades.
3. Reduir la incidència de les infeccions.
4. Optimitzar l’ús dels medicaments antimicrobians.
5. Assegurar que es realitzen les inversions sostenibles en la lluita contra la resistència als antibiòtics.
1.4. Lluita contra els superbacteris
1.4.1. ¿Tornada als antibiòtics antics o desenvolupament de nous?
Davant els creixents casos de resistència a antibiòtics, la comunitat científica d’arreu el món tracta de buscar solucions factibles i urgents. Una d’aquestes és la re-avaluació de l’ús de compostos antibiòtics que s’usaven amb anterioritat però substituïts després perquè podien resultar menys efectius i/o causar efectes secundaris derivats de la seva toxicitat (15).
A causa del baix nivell d'ús de molts dels vells antibiòtics, aquests s'han mantingut actius contra un gran nombre dels fonamentals agents bacterians que causen malalties en l’actualitat. Per això s’està considerant tornar a incloure’ls en els tractament clínics.
Es poden destacar les polimixines, la fosfomicina, l’àcid fusídic, el clotrimazol, els aminoglicòsids i el cloramfenicol com una alternativa vàlida per al tractament d'infeccions difícils de tractar. La disponibilitat de nous mètodes de modificació bioquímica, genètica i molecular ofereix una esperança als inconvenients de toxicitat i d'eficàcia d'aquests agents.
Per contra, sempre existeix la possibilitat d’iniciar la recerca o síntesi de noves substàncies bactericides que, pel fet de no haver estat mai exposades als bacteris, hi hauria una major possibilitat de que els bacteris no presentessin cap resistència i, per tant, s’ assegurés la seva mort. No obstant, també s’hauria de tindre en compte el cost econòmic i el temps invertit que això suposa.
El perill que segueix existent amb l’ús d’aquestes dues opcions és que, si es repeteix l’aplicació i l’ús inadequats d’aquests antibiòtics, es poden repetir els casos d’adquisició de multiresistències.
1.4.2. Teràpia fàgica
1.4.2.1. ¿Què són els bacteriòfags o fags?
Els bacteriòfags són un tipus de virus amb capacitat de parasitar exclusivament els procariotes (Figura 9). De fet, la seva capacitat predadora de fags és tan efectiva que, segons estimacions recents, cada 48 hores la meitat de la població bacteriana mundial és destruïda pels fags. Han estat el focus de molts estudis genètics i, tot i que la vasta biodiversitat de bacteriòfags segueix sent majoritàriament desconeguda (s’estima que sols el 0.0002% de material genètic ha estat enregistrat, ja siga DNA, RNA ambdós de doble o simple cadena (16, 17)), se’n poden oferir una gran varietat d’aplicacions mèdiques. Una d’aquestes és la utilització dels bacteriòfags com a agents antibiòtics. La clau de l’èxit d’aquests éssers roman en la seva activitat lítica.
Des de temps remots s’han documentat casos d’aigües provinents de rius amb capacitat de sanar malalties miraculosament. Segles després, gràcies al desenvolupament progressiu de la ciència, es formularen les primeres hipòtesis sobre les possibles causes d’aquestes propietats curatives. Un exemple es pot trobar al riu Ganges quan, el 1896, Ernerst Hanking hi identificà algun tipus de substància amb activitat antibacteriana contra Vibrio cholerae (22). El 1910, Felix d’Herelle, després de diversos estudis de control microbiològics, observà per primera vegada virus capaços de destruir bacteris i els va batejar així amb el nom de “bacteriòfags” o “virus menjadors de bacteris” (23, 24, 25). Anys després va provar la seva eficàcia contra infeccions amb pacients humans. Tot i això, el descobriment de la penicil·lina i posterior desenvolupament d’antibiòtics per part de la indústria farmacèutica, relegà a un segon pla l’estudi dels fags amb propòsits clínics fins quasi l’oblit total (sols uns pocs països de l’Europa de l’Est continuaren amb la recerca i fundaren instituts centrats en tals propòsits, entre els quals destaca el Institut Hirszfield d’Immunologia i Teràpia Experimental). En resum, es pot afirmar, doncs, que l’ús de fags per combatre malalties infeccioses no resulta ser un mètode “innovador”, però redescobrir-lo podria resultar un dels remeis a la crisi de resistència antibiòtica a la qual s’enfronta la nostra societat.
Hui en dia, s’anomena teràpia fàgica o fagoteràpia a l’ús terapèutic de fags específics amb cicle lític per tal de tractar infeccions provocant la mort dels bacteris patògens (18). Però, ¿com ho aconsegueixen els virus? El model general del fag més estudiat fins llavors, el fag T4, pot explicar aquest procés.
T4 és infectiu de E. coli i presenta un cicle vital exclusivament lític. La partícula viral està constituïda per una càpsida on emmagatzema el DNA de doble cadena, un collar i una cua on resideixen les fibres responsables de l’adhesió del fag a la cèl·lula que infecta. Quan el DNA víric penetra la membrana cel·lular de l’hoste és capaç de controlar la seva maquinària de síntesi proteica mitjançant l’expressió de tres grups diferents de gens; gens que es transcriuen en temps diferents rere la infecció: els primerencs immediats, els primerencs tardans i els tardans. Els primerencs es transcriuen i tradueixen abans de replicar el DNA víric. Algunes de les proteïnes resultants són les responsables d’aturar la síntesi de macromolècules del bacteri hoste. Poc després de la infecció, el DNA de la cèl·lula hoste és degradat per una DNAasa codificada per un gen primerenc. Aquest enzim no actua sobre el DNA del T4 ja que no conte citosina sinó 5-hidroximetilcitosina i l’enzim no és específic d’aquest DNA. A més, alguns dels residus de 5-hidroximetilcitosina es troben glicosilats.
El DNA del fag T4 conté derivats de la citosina a causa de l’acció de varis enzims codificats pel genoma del fag i sintetitzats en la fase primerenca de la infecció. Una pirofosfatasa hidrolitza els nucleòtids dCTP en dCMP evitant-ne així la seva incorporació al DNA víric. Una hidroximetilasa transforma els dCMP en 5-hidroximetil dCMP i una quinasa els fosforila fins dCTP podent incorporar-se, ara sí, al DNA sintetitzat. Per últim, alguns residus són glicosilats per acció d’altre enzim codificat pels virus.
La síntesi de les proteïnes tardanes es dóna junt amb la replicació del DNA del T4. En aquesta etapa es sintetitzen les proteïnes que configuren les parts del virus i lisozims. Aquests últims digereixen la paret cel·lular bacteriana produint la seva ruptura i matant el bacteri, part clau de la teràpia.
1.4.2.3. Avantatges i desavantatges de l’ús clínic de bacteriòfags
Cal analitzar els pros i contres per tal de ser conscients de quines alternatives pel tractament d’infeccions són les més efectives. Amb aquesta finalitat, s’exposaran ací alguns dels arguments usats per defensar o rebutjar l’ús clínics dels bacteriòfags.
- La majoria de bacteriòfags infecten només a una espècie de bacteris o una soca determinada dins d’una espècie. Aquest tret ofereix una sèrie d’avantatges que els antibiòtics no són capaços de complir. Utilitzant un tractament basat en bacteriòfags, hom s’assegura que no sorgirà cap tipus de complicació mèdica derivada d’infeccions secundàries per l’atac de la microbiota natural del pacient. Per exemple, s’ha observat que l’abús d’antibiòtics pot ocasionar alteracions de la flora bacteriana de l’intestí gros, ocasionant que C. difficile (un bacteri oportunista que es troba formant part de la flora en petites proporcions) proliferi excessivament arribant a causar des d’una diarrea relativament benigna fins a colitis pseudomembranosa.
- D’altra banda, es palesa una vessant més interessant des del punt de vista evolutiu. L’adquisició de resistència a qualsevol medi selectiu es pot donar de forma normal en la natura (tot i que se n’està accelerant el procés de manera antropogènica). Així mateix, els bacteris també poden mostrar amb el pas del temps més fermesa contra la infecció dels fags; malgrat que ocorre amb una freqüència deu vegades menor que l’adquisició de resistència als antibiòtics. No obstant, la població vírica en contacte amb els bacteris resistents “aprèn” noves formes d’atac, és a dir, coevoluciona conjuntament. Això és possible mitjançant dos processos bàsics: mutació-selecció i recombinació.
Cal començar explicant l’equilibri existent entre mutació i selecció. La força a nivell evolutiu de les mutacions és, en general, molt dèbil i és per això que les freqüències gèniques poblacionals en un equilibri teòric estan determinades per la interacció mutació-selecció natural. La mutació recurrent i la selecció natural actuen com a forces contràries sobre els al·lels. Mentre que les mutacions introdueixen nova variabilitat genètica en les poblacions, (sempre que no es donen retromutacions amb la mateixa freqüència), la selecció natural la disminueix: elimina els individus portadors dels al·lels el fenotip resultant dels quals els confereix una menor eficàcia biològica mentre que afavoreix els de major eficàcia. Així doncs, en aquest cas, quan aparega una mutació en el genoma bacterià que els permeta sobreviure en presència d’antibiòtics o fags, aquesta es fixarà en la població. El mateix ocorrerà amb els fags; quan sorgeixa una mutació que els permeta manifestar una major eficiència d’infecció, aquesta també es fixarà en la població i el tractament podrà prosperar com estava previst. També cal nomenar, que habitualment s’usen còctels de bacteriòfags específics i antibiòtics per agilitzar el procés i dificultar, en la mesura del que és possible, que els bacteris s’immunitzen contra els fags i sobrevisquen.
S’ha parlat, també, de recombinació. Els bacteris són capaços d’intercanviar material genètic, però no són els únics. Quan fags amb diferents al·lels infecten la mateixa cèl·lula, es pot donar el fenomen de la recombinació entre els genomes vírics. Quan les noves partícules que conformen els virus es tornen a acoblar, algunes d’elles emmagatzemaran fragments de material genètic amb diferent informació que la inicial , és a dir, amb diferents combinacions al·lèliques que les inicials. Ací roman la clau de l’evolució dels fags.
- Els fags es repliquen en el lloc d’infecció, de manera que actuen de manera més ràpida que els antibiòtics que circulen des del lloc d’aplicació (via oral, cutània, intravenosa...) fins el torrent sanguini per tot el cos i, finalment, al lloc d’infecció.
- El cost de disseny de teràpies amb fags és més barat i ràpid que el desenvolupament de nous antibiòtics.
- Caldria fer estudis molt personalitzats de cada pacient sotmès al tractament. S’hauria d’analitzar quin tipus d’infecció s’ha produït, quins medicament han resultat ineficaços i fer un disseny del tractament que consistiria en combinacions específiques de bacteriòfags seleccionats i antibiòtics no usats fins el moment. Moltes vegades resulta impossible fer aquest tipus d’estudis pels recursos econòmics que això suposa.
- El tractament pot no ser eficaç segons l’estat en el qual es trobe el sistema immunològic del pacient. Cal recordar que els virus estan formats per partícules que resulten alienes al nostre organisme (antígens) i que aquest reaccionarà per tal d’eliminar-los. Estudis farmacocinètics suggeriren que els fags eren eliminats del torrent sanguini per l’acció d’ immunoglobulines o retesos a la melsa que actuaria a mode de filtre. D’altra banda, s’ha de considerar la possibilitat que es produïsquen fortes reaccions al·lèrgiques o, fins i tot, xocs anafilàctics.
- S’ha de dur un control exhaustiu per evitar l´ús de fags atemperats, és a dir, els que passen per un cicle lisogènic integrant-se temporàniament en el genoma de l’hoste. Aquests són descartats perquè no poden produir una lisi ràpida. A més, poden originar fenòmens de conversió mitjançant la transmissió de factors de virulència.
1.4.3. “Enzibiòtics”. Enzims lítics codificats pels fags
En parlar de la teràpia fàgica, s’ha especificat que la lisi dels bacteris forma una part clau del tractament. Recentment s’han dut a terme investigacions que demostren com certs enzims (en aquest cas endolisines proteiques) codificats pels bacteriòfag, poden usar-se de manera aïllada com noves eines terapèutiques contra aquelles infeccions que no poden ser tractades eficientment amb antibiòtics convencionals (20).
Rere la infecció al bacteri, els fags sintetitzen endolisines que produeixen la mort de la cèl·lula hoste des del seu interior. Les endolisines presenten dominis específics que les permeten dirigir-se i ancorar-se selectivament sobre la paret cel·lular bacteriana. Llavors, s’inicia la translocació cap a la paret i ho fan gràcies a unes proteïnes anomenades holines. Aquestes actuen perforant la membrana cel·lular i permetent a les endolisines accedir a la capa més externa del bacteri. Allí s’uneixen al principal polímer estructural, els peptidoglicans, i hidrolitzen els enllaços que l’estabilitzen, produint així una lisi hipotònica i la posterior alliberació de la progènie fàgica (Figura 10).
Figura 10.- Cicle lític d'un fag atemperat (biologiamedica.blogspot.com)
Experiments, tant in vitro com en in vivo, han demostrat l’eficiència de les endolisines per produir la mort dels bacteris.
¿Quins avantatges presenta aquest mètode front la teràpia fàgica? El seu efecte és immediat, no hi ha risc de transducció de factors de virulència i, fins llavors, no s’ha observat cap fenomen de resistència a endolisines.
Cada cop més es coneix l’enorme diversitat catalítica d’aquests enzims, la seva funció i organització a nivell atòmic, els mecanismes pels quals actuen i, fins i tot, les seqüències del material hereditari que les codifiquen. Això suposa un avantatge enorme en la biotecnologia dels aliments per la creació d’organismes modificats genèticament capaços de codificar les seves pròpies endolisines contra els bacteris pels quals són normalment infectats.
1.4.4. Reversió de la resistència bacteriana
Fins ara les solucions s’han centrat en nous mecanismes per causar la mort directa dels superbacteris. Però també resulta interessant explorar una altra vessant: restaurar l’eficiència dels tractaments revertint els bacteris al seu estat més susceptible a l’acció dels antibiòtics. Per a això és interessant endinsar-se en l’enginyeria genètica (26).
1.4.4.1. Un clar exemple
Per poder explicar amb claredat en què consisteix aquesta proposta, s’ha tingut en compte un article que l’exemplifica perfectament. L’escrit es publicà al 2011 i provava l’eficàcia d’un sistema que cessa la resistència antibiòtica de l’estreptomicina i l’àcid nalidíxic causada per mutacions en els gens rpsL i gyrA, respectivament (27).
L’estreptomicina és un bon antibiòtic contra els bacteris gram-negatius i gram-positius; no obstant, s’ha observat com M. tuberculosis, causant de la tuberculosi, ha desenvolupat resistència. El 24% - 85.2% dels bacteris resistents presenten mutacions al gen rpsL, el producte del qual és una proteïna altament conservada de la subunitat petita 30S del ribosoma. Les mutacions provoquen que es codifique una proteïna de conformació diferent que evita la unió inhibitòria de l’estreptomicina per la qual cosa la proteïna continua essent funcional i la cèl·lula sobreviu. A més, se sap que són recessives respecte l’al·lel salvatge del gen (wt-rpsL). Per tant, és lògic pensar que si s’aconseguira proporcionar al bacteri una còpia de l’al·lel dominant, l’estreptomicina tornaria a ser eficient.
En aquesta direcció es dirigí l’estudi. Es simularen les condicions anteriors en dues soques d’E.coli amb dues mutacions diferents al gen rpsL i es transformaren usant dos mètodes: plasmidis artificials i fags modificats genèticament, ambdós - dissenyats específicament per evitar recombinació homòloga indesitjable entre l’inserit i el cromosoma bacterià-, i portadors del gen d’interès i un gen marcador o “reporter” (selecció dels bacteris transformats en un medi selectiu) de resistència a la tel·lurita.
Per comprovar si a la pràctica el gen introduït retornava la sensibilitat a l’antibiòtic, es va determinar el MIC de les soques abans i després de la transformació. MIC significa “Minimum Inhibitory Concentration” i fa referència a la mínima concentració d’antibiòtic requerida per inhibir el creixement visible dels microorganismes. Els resultats que s’obtingueren foren els següents.
Per la transformació amb el plasmidi:
- soca Sm6. MIC inicial i plasmidi control: 100g/ml. MIC final: 25g/ml.
- soca Sm13. MIC inicial: 200g/ml. MIC final: 6,25g/ml.
La dràstica reducció del MIC confirma l’eficiència del mètode.
Per a la lisogenització pel fag:
- soca Sm13. MIC inicial i fag control: 200g/ml. MIC final: 50g/ml.
Se sospitava que la menor disminució de la sensibilització podria ser causada al nombre de còpies introduïdes al fag. En augmentar-ne el nombre el MIC passà a ser de 1,56g/ml.
Es repetí l’experiment per l’àcid nalidíxic, un antibiòtic de la família de les quinolones. Les quinolones actuen sobre l’enzim girasa, codificada pel gen gyrA (wt-gyrA també dominant), aturant la replicació del DNA. Les mutacions en el gyrA són observades en una regió anomenada QRDR (regió determinant de resistència a les quinolones).
La sensibilització tornà a aparèixer tant amb l’ús de plasmidis com amb fags.
Coneixent aquestes dades, els autors de l’article plantegen un sistema de dispersió des fags modificats en les instal·lacions mèdiques (resulten un millor vehicle de transmissió dels gens d’interès que els plasmidis i són més segurs). En contrast amb l’ús convencional dels antibiòtics i la teràpia fàgica, aquesta proposta canviaria potencialment la naturalesa de les infeccions nosocomials en direcció a ser cada cop més susceptibles als antibiòtics en lloc de més resistents.
1.4.4.2. Avantatges i desavantatges
- S’utilitzen tècniques d’enginyeria genètica per a la construcció dels fags són senzilles. S’abarateix el cost.
- La coevolució ha fet possible que trobem fags específics de qualsevol bacteri.
- Les tècniques usades per evitar la recombinació homòloga són eficaces i també senzilles. Amb les eines de les que es disposen hui en dia, és pot aconseguir per a qualsevol gen que es desitge.
- El cassette gènic podria incloure altres gens de sensibilització de manera que disminueix la probabilitat d’aparició de bacteris resistents a diversos antibiòtics simultàniament. A més, aquests gens solen tindre un caràcter dominant sobre les mutacions que confereixen resistència en una àmplia varietat d’espècies de bacteris. La reversió s’aconsegueix amb mínim una còpia del gen salvatge.
- Es podrien dissenyar en un futur constructes més segurs. El constructe pot ser sintetitzat de manera que la presència del cassette gènic siga essencial per a que el gen de resistència siga funcional. Aquest es podria situar, sense promotor, al darrera del cassette; la seva expressió dependria de l’expressió dels primers.
- L’aplicació dels fags a les superfícies de les instal·lacions hospitalàries també és senzilla. S’abarateix el cost.
- S’evita l’ús dels fags dins el cos dels pacients, per la qual cosa se superen els problemes derivats de possibles rebutjos pel sistema immunològic, reaccions de toxicitat…
- El cicle lític dels fags sols s’indueix a temperatures elevades. Si els bacteris infecten els humans o altres animals homeoterms (“de sang calenta”) que presenten en eixe moment una temperatura corporal alta, els fags entrarien en el cicle lític causant la mort directa dels bacteris.
- La utilització dels fags als hospitals canviaria la naturalesa de les poblacions que constitueixen la primera causa de malalties nosocomials. L’àmplia transferència de cassettes gènics augmentaria la població dels bacteris tractables amb antibiòtics i desplaçaria el nínxol ecològic que ocupen els bacteris resistents.
- Per ara aquestes aplicacions es limiten a l’ús en hospitals, però no es descarta que es puga ampliar a d’altres indrets.
2. Conclusions
És necessari remarcar alguns aspectes que es poden considerar especialment importants.
Cal llegir entre línies per entendre fins on arriba la magnitud del problema al que s’enfronten les nostres societats. En primer lloc, el mateix fet de batejar aquests bacteris com a superbacteris és, en sí, preocupant. La pròpia existència d’uns éssers patògens que són resistents a mètodes, en principi, quasi infal·libles, i pels quals no existeix un mètode d’eradicació perfecte, suposa una amenaça equiparable a altres catàstrofes de nivell mundial com el canvi climàtic.
El segon lloc del nostre top de punts importants l’ocupa el caràcter mundial que adquireix aquest problema; fins i tot l’OMS ha iniciat un pla d’acció per aconseguir aturar l’emergència de noves soques bacterians resistents a diversos antibiòtics. Sabem que la globalització juga un paper fonamental a favor de l’augment dels Superbacteris. A mesura que augmenten les xarxes de transport internacionals i, fins i tot, intercontinentals, es facilita la dispersió de xicotets elements naturals, com puguen ser les espores o també els bacteris. Per tant, no és d’estranyar que en poc temps s’haja magnificat el problema. Així doncs, aquesta és una amenaça a nivell mundial que afecta a tots per igual; països considerats grans potències mundials, països en vies de desenvolupament, rics, pobres, homes, dones, ancians, nens...
És també important analitzar on radica l’arrel del problema. Hem arribat fins a aquest punt per haver fet un mal ús dels antibiòtics (com ja hem dit: dosi mal calculades, malalties no provocades per bacteris tractades amb antibiòtics, mal calculat el temps de pressa d’antibiòtics...). La “culpa” no la tenen els bacteris, perquè ells han evolucionat com ho farien a la natura adaptant-se a unes condicions extremes o limitants per a ells; ha estat l’ésser humà qui, buscant l’utòpic món sense malalties, ha creat aquests monstres. Així, les mesures que hem de prendre per eradicar aquest problema no són tan sols científiques i mèdiques; ha d’haver-hi una conscienciació i una educació a nivell mundial.
Per últim, cal remarcar la complexitat de la situació. No existeix una única forma de resistència antibiòtica com tampoc existeix una única forma de combatre-la. És molt possible que tornar a una època en la que els antibiòtics eren medicaments eficaços i segurs coste molts anys i diners de solucionar. Hem estat completament cecs davant aquesta Amenaça Fantasma.



 Figura 1.- Alexander Fleming al seu laboratori.
Figura 1.- Alexander Fleming al seu laboratori.
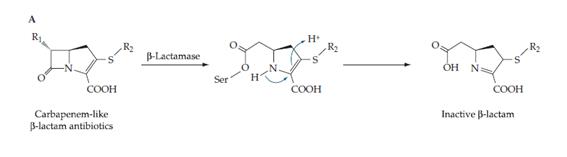 Figura 2.- La serina de les B-lactamases que contenen aquest aminoàcid, forma un enllaç covalent amb l'anell dels B-lactams.
Figura 2.- La serina de les B-lactamases que contenen aquest aminoàcid, forma un enllaç covalent amb l'anell dels B-lactams.
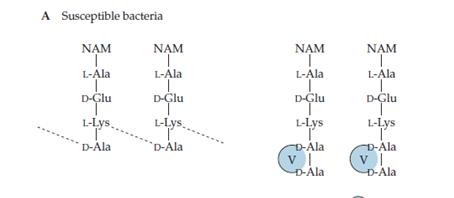 Figura 3.- Mecanisme d'acció de la vancomicina en bacteris sense resistència. (Wilson, BA et al. 2007)
Figura 3.- Mecanisme d'acció de la vancomicina en bacteris sense resistència. (Wilson, BA et al. 2007)
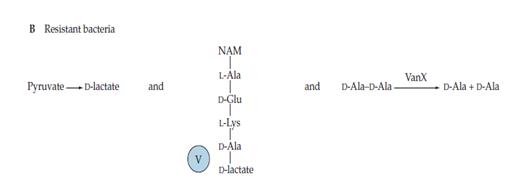 Figura 4.- Mecanisme bacterià per resistir a la vancomicina. (Wilson, BA et al. 2007).
Figura 4.- Mecanisme bacterià per resistir a la vancomicina. (Wilson, BA et al. 2007).
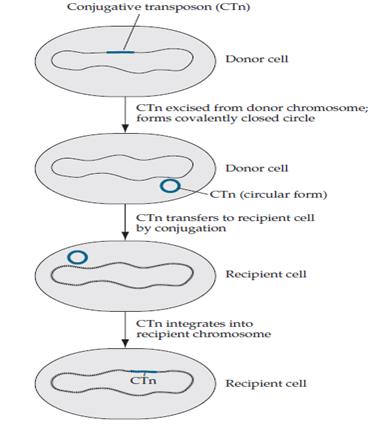 Figura 5.- Transferència d'un transposó (Wilson, BA, 2007).
Figura 5.- Transferència d'un transposó (Wilson, BA, 2007).
 Figura 6.- Retrat d'Ignác Fülöp Semmelweis (chronotopeblog.com)
Figura 6.- Retrat d'Ignác Fülöp Semmelweis (chronotopeblog.com)
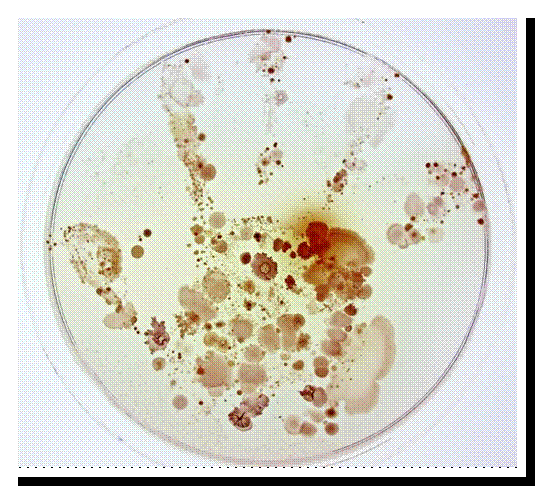 Figura 7.- Fotografia guanyadora del primer premi de la 13a edició del seu certament Fotociencia. Empremta d'una xiqueta de 6 anys després de posar la seua mà sobre una placa de Petri.
Figura 7.- Fotografia guanyadora del primer premi de la 13a edició del seu certament Fotociencia. Empremta d'una xiqueta de 6 anys després de posar la seua mà sobre una placa de Petri.
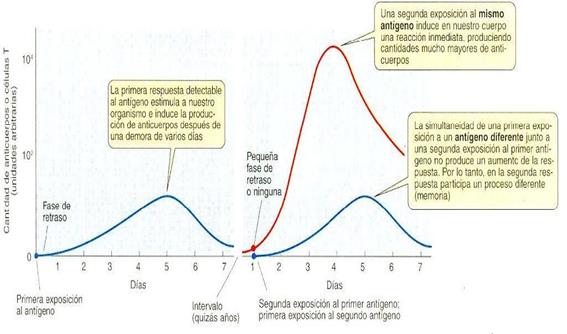 Figura 8.- Resposta immunològica primària i secundària (epideiologiamolecular.com)
Figura 8.- Resposta immunològica primària i secundària (epideiologiamolecular.com)
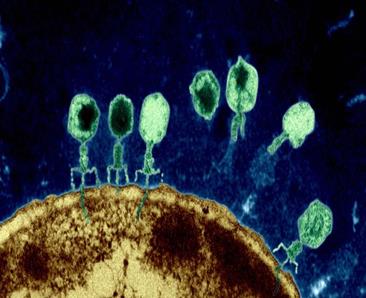 Figura 9.- Infecció per fags (es.wikipedia.org)
Figura 9.- Infecció per fags (es.wikipedia.org)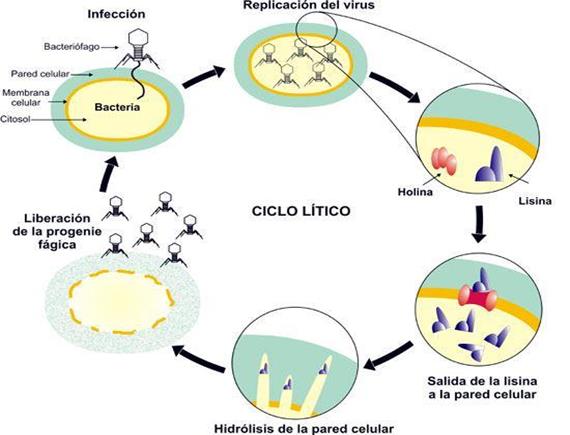 Figura 10.- Cicle lític d'un fag atemperat (biologiamedica.blogspot.com)
Figura 10.- Cicle lític d'un fag atemperat (biologiamedica.blogspot.com)